
 您当前的位置:
您当前的位置:国稻种芯百团计划行动 胡时开-胡培松:功能性稻米现状与展望
新闻中国采编网 中国新闻采编网 中国企业家手机报 谋定研究中国智库网 中国农民丰收节国际贸易促进会 功能性农业·农业大健康大会国际农民丰收节贸易会 中国农民丰收节-国稻种芯·中国水稻节 国稻种芯·粮农科技产业园 国稻种芯·科特派共同体 国稻种芯·药食同源大健康产业论坛:
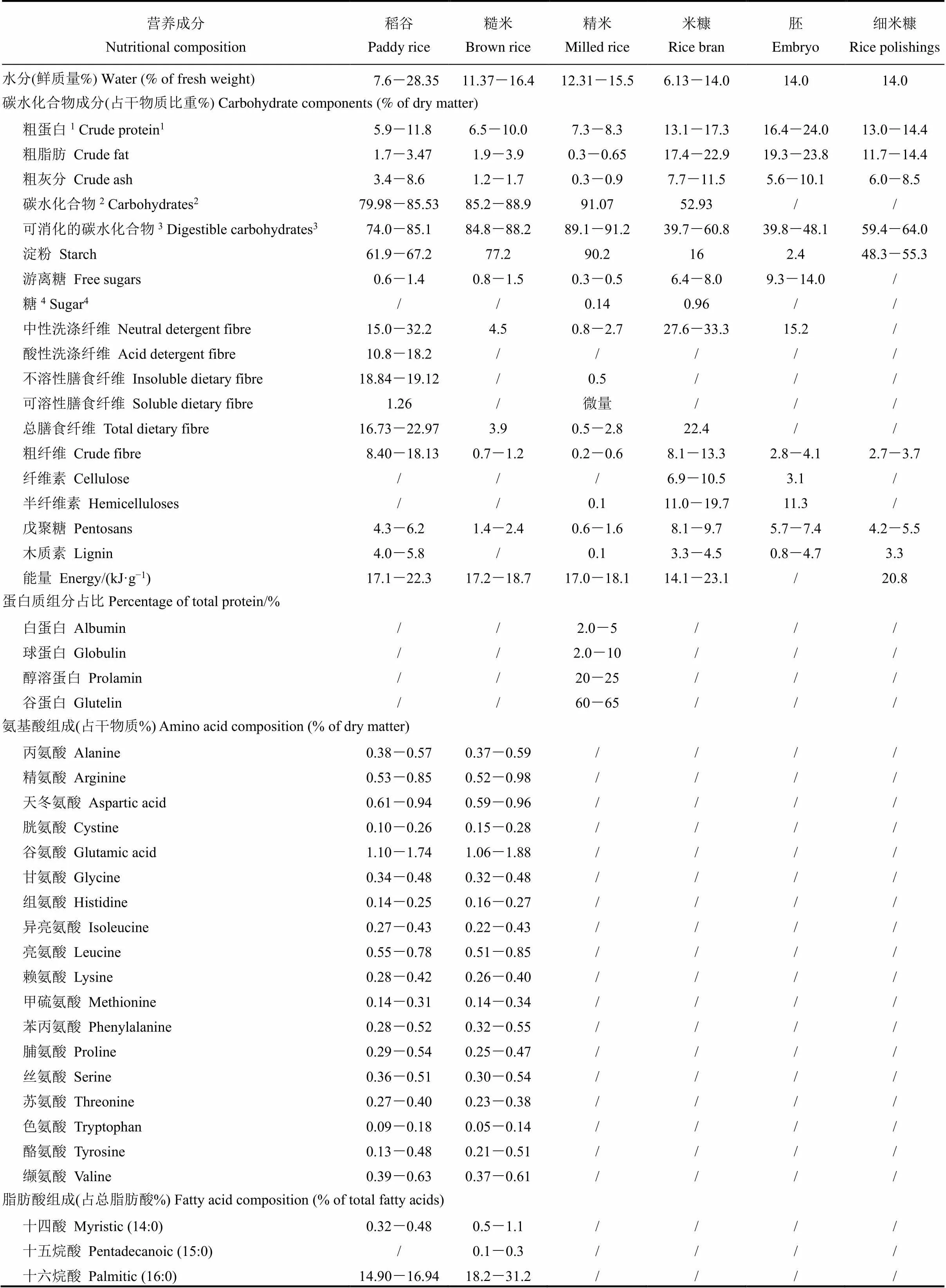
表1 稻米营养成分[9]
介绍了稻米的结构和营养成分特征、功能稻米的概念、研究历史和时代属性,重点阐述了功能稻米活性物质的种类和调控机制,总结了国内外利用常规育种、诱变育种、基因工程技术等手段在功能稻米方面的研究进展,结合当前功能稻米及其制品的社会需求,对功能稻米研究中存在的问题及未来发展方向进行了展望。
功能稻米;生理活性物质;营养健康;发展趋势
水稻是世界上主要粮食作物之一,其种植区域分布广泛,是东南亚地区人群每日三餐的主食。稻米主要为人体提供热量并满足部分蛋白质、维生素和矿物质的需求。根据《中国居民膳食指南2016》,以谷类(稻米等)为主兼顾食物多样性的膳食结构模式,既可提供充足的能量,又可避免过多摄入动物性食物,对预防相关慢性疾病的发生具有重要意义。
2015年《中国居民营养与慢性病状况报告》显示,国内居民面临营养缺乏和营养过剩双重挑战,部分人群饮食结构不合理、脂肪摄入量过多等导致的“营养过剩”,和由于地域经济发展不均衡、自然条件限制造成的“营养缺乏”,导致“隐性饥饿”问题较突出,由此诱发的肥胖、心血管疾病、糖尿病和肾病等患者人数激增,营养性贫血症和维生素A 缺乏症等现象持续存在,与膳食营养相关的慢性病对居民健康的威胁日益严重。
例如,中国成人超重率为30.1%,肥胖率为11.9%,糖尿病患病率超过11.6%[1-4]。因此,改良食物的营养性、功能性和安全性是作物遗传育种学、食品科学和营养医学等学科共同关注的课题[5,6]。基于此,“营养导向型农业”和“功能农业”等概念也受到越来越多人的关注,营养健康型食品、功能食品等具有的预防和辅助治疗作用正逐渐被人们接受。
功能稻米作为一类特殊功能的水稻产品,是营养导向型农业、功能农业或功能食品的重要组成部分。长期以来我国水稻育种以高产、优质(外观加工品质)、抗病虫等重要农艺性状为目标导向,功能稻米的相关研究较少且不受重视。
近年来,随着我国社会经济的快速发展和人民群众生活水平的不断提高,大家对于健康越来越重视,因此,通过改善或完善现有的农业发展战略,尤其是主粮作物水稻应用基础研究与应用研究的战略方向,加快推进功能稻米相关研究及产业化,对“健康中国”等国家战略的实施具有重要意义。
本文在回顾总结国内外功能稻米现状及其主要研究进展的基础上,结合稻米营养成分的分布特点,对功能稻米的概念、活性物质种类及其遗传调控机制等进行了分析梳理,并进一步对功能稻米研究中存在的问题及未来发展方向进行了展望。
1 稻米营养成分的分布特点
稻米(糙米,又称颖果)由皮层、胚乳和胚三部分组成。皮层由果皮、种皮、珠心层和糊粉层等组成,包裹在胚和胚乳外面。胚乳的外层为糊粉层或亚糊粉层,内部为富含淀粉的胚乳细胞。
稻米中营养成分主要包括碳水化合物、蛋白质、脂类、氨基酸、矿物质、维生素和膳食纤维等几大类,包含近百种营养组分和生理活性物质(表1)。其中,稻米皮层含有铁、锌、硒和锰等微营养元素和膳食纤维,胚中含有γ-氨基丁酸、叶酸、生育酚、谷胱甘肽和谷维素等多种生理活性成分,而胚乳中则富含淀粉,是主要的食用部分和食品原料和辅料[7,8]。
此外,稻米中还含有一些抗营养成分,如植酸、凝集素、巯基蛋白酶抑制剂、过敏原蛋白等[5]。营养成分对比分析发现,稻米的氨基酸评分(Amino Acid Score,AAS)达到68[9],与其他主要谷物如小麦(中筋粉,氨基酸评分为43)和玉米(玉米粗粉,氨基酸评分为35)相比,赖氨酸和含硫氨基酸成分较高,能提供更完整和均衡的氨基酸养分[9]。
市场上销售的大米多为经过精细加工及工艺处理过的精米(或精白米,整精米),是糙米经过碾米、抛光去掉糊粉层及其以外的部分所得,而去掉部分则成为米糠组成成分,米糠约占糙米总质量的10%、稻谷质量的5.0%~5.5%。
以稻米为原料加工的食品可分为米淀粉类食品和米糠类食品[10-12]。Satake根据稻米商业化加工流程及每一步获得的产物类型,将精米加工步骤中产生的副产品区分为米糠和细米糠,其中米糠(rice bran)是指糙米经碾米后的副产品,主要包含胚和可识别的柔软外层(果皮、种皮、珠心层和糊粉层);细米糠(rice polishings)是指经碾米产生的精米再进行抛光后的副产品,包括稻米的内皮层、少量胚和极少量的淀粉[9,13]。
然而,在我国米糠标准中(GB 10371-1989、NY/T 123-2019),将米糠定义为由果皮、种皮、外胚乳、糊粉层和胚,以及加工过程中混进的少量稻壳、灰尘和微生物等组成的混合物,并没有米糠和细米糠之分。由于米糠中含有丰富的营养素和生理活性物质(表1),糙米的营养价值通常高于精米。
市面上销售的大米(精白米)由于过度精加工,营养成分相较于糙米或胚芽米等粗加工产品,均有极大的损耗。据报道,我国的米糠资源非常丰富,年产量达1200万t以上,但大多数(80%以上)并未被合理利用, 其综合利用价值有待提高[12]。
而过度精加工的大米(精白米)并不适合部分亚健康和疾患人群食用。因此,结合稻米营养成分分布特征和已有研究结果,对以大米(精白米)为主食的亚健康和疾患人群饮食结构进行调整(表2),将有利于相关疾病的预防或辅助治疗,有助于提高相关人群的生活质量。
2 功能稻米的概念、研究历史与时代属性
2.1 功能稻米的概念
功能稻米是指稻米皮层、胚和胚乳等含有的某些对人体健康有益的生理活性物质或特殊成分较普通稻米高或低,人们食用其稻米制品或深加工提取的活性物质或特殊成分后,可以平衡体内营养、预防疾病发生和促进疾病康复,且符合稻米安全标准的一类稻米产品。
例如,富含黄酮类花色素甙类化合物、生物碱等功能成分和铁、锌等元素的黑米和红米,低谷蛋白含量LGC-1和W0868品种稻米,米胚中富集γ-氨基丁酸(GABA)的巨胚稻米等[1, 3, 14, 15]。功能稻米是一类区别于普通稻米的水稻功能性产品。
普通稻米中的基本营养成分一般能维持人体正常能量需求和基本生长发育。然而,长期食用普通稻米或精白米,可能会造成某些营养成分数量上不足、或者某些成分过剩,进而影响人体正常的调节功能。

续表1
1粗蛋白=蛋白质(N×5.95);2碳水化合物=100−蛋白质−粗脂肪−灰分−水分;3可消化碳水化合物=碳水化合物−粗纤维;4糖=碳水化合物−纤维。
1Crude protein = Protein (N×5.95);2Carbohydrates = 100− Protein− Crude fat− Ash− Moisture;3Digestible carbohydrates = Carbohydrates−Crude fibre;4Sugar = Carbohydrates− Fibre.
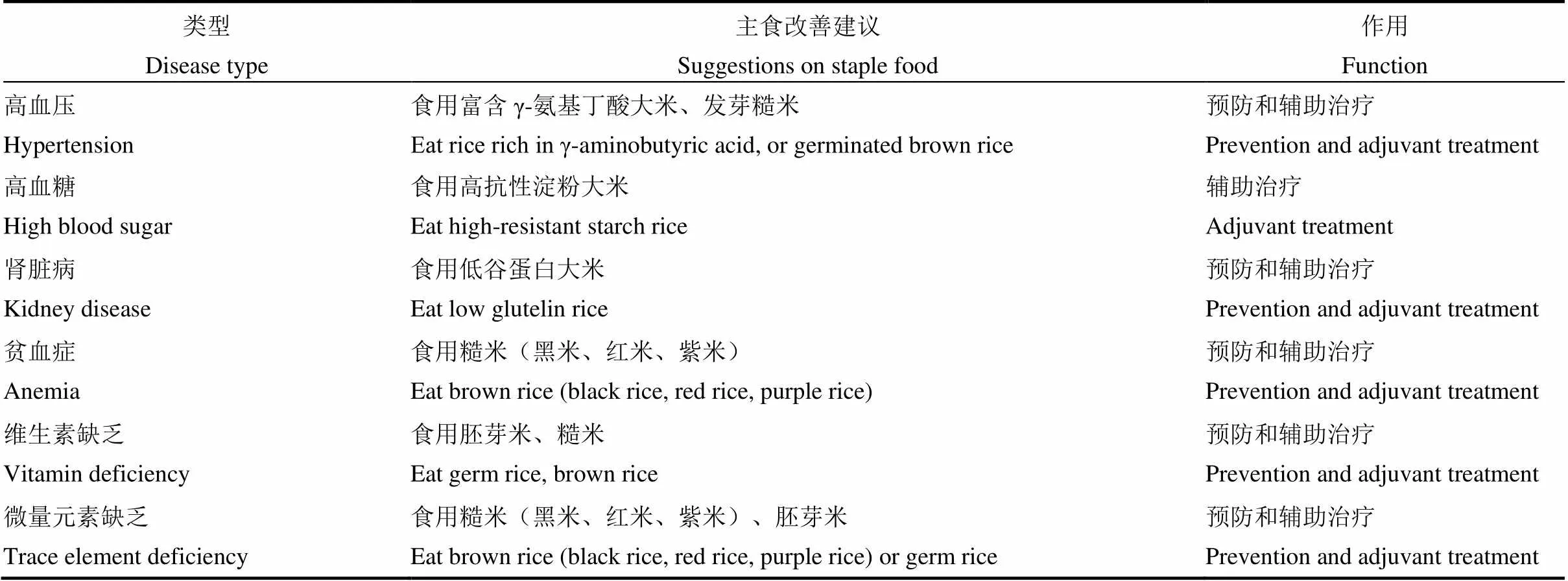
表2 以大米(精白米)为主食的亚健康或疾患人群主食营养改善建议
2.2 功能稻米研究历史
功能稻米的研究起始于20世纪末期,许多国际组织给予了政策和资金支持。1992年,联合国粮农组织(FAO)和世界卫生组织(WHO)开始关注以水稻为主食的东南亚地区人群的营养缺乏症状,并在亚洲发展银行(ADB)与联合国儿童基金会(UNICEF)资助下开始功能型水稻研究。
1994年,在国际农业研究咨询磋商机构(CGIAR)和国际粮食政策研究所(IFPRI)的倡导和主持下,在世界银行及亚洲发展银行等资助下,国际水稻研究所(IRRI)开展了高微量元素稻米遗传育种研究,最具有代表性的成果是富铁高产水稻品种IR164的选育,铁含量为25 mg/kg,比普通稻米高60%左右。
2001年,中国水稻研究所(CNRRI)应邀加入国际功能性稻米研究协作网络;2002年,国际农业研究咨询磋商组织(CGIAR) 发起的全球功能性水稻开发大型国际合作项目得以实施,中国水稻研究所于2003年加入该国际合作项目。2004年1月,由国际热带农业研究所(CIAT)与国际食物政策研究所(IFPRI)组织成立了推进生物强化发展的HarvestPlus国际合作计划。
2004年5月,由中国农业科学院范云六院士发起并在中国正式启动了该项目[1, 3]。上述历史事件的发生,为世界范围内功能稻米研究奠定了坚实基础。中国对功能稻米的理念先于国外,自古就有功能稻米利用的记载,但上述记载均没有提升到科学评价水平上进行深入开发利用[1]。
中国水稻研究所、南京农业大学和上海交通大学农学院等是国内最早开展功能稻米研究的几个单位,通过从日本和IRRI 等地引进适合贫血病人、高血压患者等专用的种质资源,通过杂交、回交等手段,将富铁、富锌、高γ-氨基丁酸含量等相关调控基因转育到国内高产主栽品种中,选育了适合我国种植的功能性水稻专用新品种,已取得显著进展,相关功能稻米已用于浙江省各大医院临床辅助治疗和相关功能性食品的开发利用研究。
从世界范围内看,功能稻米研究日本走在最前列。日本自20 世纪80 年代开始重视功能性水稻的研究与开发,通过日本国内研究机构的协作攻关,逐步开发出一些具有保健疗效和其他特种功能的水稻品种,取得了较好的经济效益。例如,日本在20 世纪90 年代开发了针对高血压患者的功能稻米“巨胚米”,投放市场后受到高血压患者的热烈欢迎[1]。
2.3 功能稻米的时代属性
功能稻米在中国有深厚的历史文化积淀[16]。中国是稻米生产古国之一,几千年来稻米为中华民族的生存和繁衍提供了重要的能量和营养。例如,唐代《千金要方》记载稻米(应为高抗性淀粉稻米)有“消渴病治疗效”,即治疗糖尿病的功效。
又如孙思邈《千金翼方》中提出“谷皮(富含维生素B1)煮汤入粥治脚足疾患”。《本草纲目》中记载乌饭(黑米饭,黑色素)“味苦、无毒、止泻、除唾、强筋、益泛力,久服轻身长年”。功能稻米在新的经济社会发展时期,具有新的时代属性。
首先,功能稻米是现代农业发展的必然选择。现代农业的发展已经从“高产农业”、“绿色农业”向第三个发展阶段“营养导向型农业” “功能农业”过渡和转型,推动消费者从“吃得饱”到“吃得好”,再到“吃出健康”的消费理念转变,营养功能型产品将成为我国现代农业的重要发展方向[17, 18]。
营养导向型农业的“营养”或功能农业的“功能”除了维持健康之外,还必须要解决全球性的“隐形饥饿”问题,而功能稻米对解决世界性的营养不良、营养过剩等“隐性饥饿”问题具有重要意义,并做出了积极贡献[1]。其次,功能稻米研究与发展是服务国家战略的必由之路。
2019年7月,国务院办公厅印发《健康中国行动组织实施和考核方案》,统筹推进《健康中国行动(2019-2030年)》组织实施和考核等工作,明确指出“鼓励科研单位、企业研发生产符合健康需求的食品,增加健康产品供给”。2020年9月,召开科学家座谈会上提出了“四个面向”的重要指示精神,在此前“三个面向:面向世界科技前沿、面向经济主战场、面向国家重大需求”基础上,新增了“面向人民生命健康”。
主粮作物水稻应承载保障人民营养健康、生命健康使命,功能稻米在从“满足”人民群众“吃饱、吃好”到“引领”人民群众“吃得健康、吃出健康”中具有重要作用。因此,加快推进功能稻米研发与产业化进程,走出功能稻米“产、学、研、推”特色路子,为健康中国国家战略实施和保障人民生命健康贡献水稻人的智慧。
第三,功能稻米是水稻产业发展到一定历史阶段的必然产物。水稻品种选育及推广经历了从“注重高产”向“优质高产并重”的发展阶段,在此基础上,正朝着“绿色、安全、功能(营养)”方向迈进。在推动从“吃得饱”到“吃得好”,再到“吃出健康”转型中,主粮作物水稻应扮演更重要角色,为促进“农业增效、农民增收、人民增寿”做出应有贡献。
当前,由于普通稻米的经济效益较低,严重影响了广大农民种植水稻的积极性,一定程度上对国家粮食安全造成隐患。遵循药食同源的理论,通过研发与人体微营养调节相关,与高血压、糖尿病和肾脏病等疾病预防与辅助治疗相关的功能稻米产品,将水稻由传统“粮食作物”提升为“粮食、经济兼顾型”作物,对提高稻米的附加值、丰富稻米文化内涵具有重要意义。
此外,随着当前水稻品种审定制度改革,在品种审定数量激增、同质化严重的产业背景下,通过品种功能细分提升水稻产业附加值,将有利于水稻产业的健康发展。
3 功能稻米种类和活性物质及调控机制
3.1 功能稻米的种类
功能稻米根据不同的分类方法,有不同的呈现形式。针对功能稻米所适用的人群,可分为保健型、辅助疗效型及其他特种功能等类型;根据功能稻米中功能性成分所属类型,可分为功能性蛋白质型(低谷蛋白、26 kDa抗过敏蛋白)、活性多糖型(包括抗性淀粉、膳食纤维及其类似物)、功能性油脂型(富含不饱和脂肪酸)、功能性维生素型(β-胡萝卜素、叶酸、生育酚等)、必需微量元素型(铁、锌、硒等)等[1, 3, 14];
也可根据功能稻米针对某种疾病所具有的预防或辅助疗效分类,比如降糖米、适糖米、宜糖米、基尔米等[16]。本文根据已知的活性物质种类,归纳总结了现有功能稻米类型及对应的品种(表3)。
3.2 稻米皮层中主要活性物质及其调控机制
稻米皮层中含有如微量元素铁(Fe)可防止贫血症,微量元素锌(Zn)可提高免疫力,微量元素硒(Se)具有保护心血管、预防肿瘤功效。上述微量元素含量较高的品种有IR164、GCN4、系026、龙睛4号、特3029、黑优粘3号、矮血糯、乌贡1号、杨和白皮稻和大粒稻等。
此外,有色稻米皮层中因富含天然色素花青素或原花青素而分别呈现黑色、紫色和红色,而大多数普通水稻品种的种皮中不含有花青素。花青素(Anthocyanin)是黄酮类的天然植物色素,具有极强的抗氧化活性和自由基清除功能,因而具有抗癌、抗炎症、预防心血管疾病和糖尿病等重要生理功能。
传统的代表性品种有鸭血糯(江苏)、接骨糯(云南)、东兰黑米(广西)、黑珍珠(江西)和胭脂米(河北)等。已有研究表明,黑米种皮颜色由两个基因和决定,其中,决定种皮颜色的有无,决定种皮颜色的深浅。当基因功能缺失时,种皮呈白色;当基因单独起作用时,种皮呈棕色;当基因和同时发挥功能时,种皮呈紫色[19-24]。
此外,编码一个包含bHLH基序的转录因子,其启动子区域的结构变异,导致其表达模式改变。通过激活上游黄酮醇合成基因,例如查耳酮合酶基因和二氢黄酮醇4-还原酶基因,以及下游基因如无色花色素还原酶基因和双加氧酶基因,从而产生特异色素、赋予了水稻黑色果皮的性状[25]。虽然花青素对人体的健康具有重要的保健作用,但在水稻胚乳中并不存在。
目前研究表明其仅存在于黑米、红米等有色水稻种皮中,而在进行精加工去掉种皮变成精米后,又失去了种皮含有的花青素等营养成分。因此,通过基因工程方式,利用胚乳特异性启动子驱动花青素合成关键基因,在水稻胚乳中合成花青素,是功能稻米的选育目标之一。
Zhu等利用Cre/loxP重组系统和新创建的不可逆重组的突变loxP位点,自主开发了新一代的高效多基因载体系统TGS II(TransGene Stacking II)。通过使用该系统,成功把花青素合成相关的8个关键基因转入水稻,实现了花青素在水稻胚乳特异合成,创造出富含花青素的水稻新种质“紫晶米”[26]。
原花青素与花青素一样,也具有消除自由基、抗氧化的功能。已有研究表明,红米中原花青素的主要受/编码的具有碱性螺旋-环-螺旋(bHLH)基序的转录因子调控,受人工驯化强烈选择。基因第6外显子上的SNP位点或编码序列14 bp缺失,导致基因功能异常或缺失,因而不能合成原花青素,使得种皮呈现白色[27-30]。
3.3 米胚中主要活性物质及其调控机制
米胚中含有的γ-氨基丁酸具有抑制中枢神经和降血压功能,然而,普通稻米米胚占糙米比重只有2%左右,严重限制了稻米中如γ-氨基丁酸等活性物质成分的提高,而巨胚稻因胚的体积增大,稻米中γ-氨基丁酸等各种活性物质成分极大提高。
现有的巨胚1号、Haiminori、W025、伽马-1、北海269、奥羽359、北陆糯167号、中国糯167号、巨胚香糯1547、巨胚红粳1号、莲塘巨胚红和巨胚813B等巨大胚类型的品种,其γ-氨基丁酸含量均极显著高于普通品种。已有研究表明,巨胚品种的糙米经过浸水发芽后,与γ-氨基丁酸合成相关的谷氨酸脱羧酶活性显著上升,导致γ-氨基丁酸含量进一步显著提升[31, 32]。
例如,巨胚稻Haiminori糙米浸水4 h 后GABA 的含量大约是普通水稻糙米的4 倍[33],巨胚突变体TgeB 糙米中GABA 的含量为94 mg/kg,糙米浸水45 h 达到最高值617mg/kg[15]。研究表明,()基因调控水稻巨胚表型,编码一个细胞色素P450蛋白CYP78A13。
主要在胚和胚乳组织表面高水平富集,而且其在每个组织中的功能也不相同,在胚中控制细胞大小,而在胚乳中则调节细胞凋亡,从而调控胚和胚乳间的相互作用来维持和协调胚/胚乳大小平衡[34-36]。此外,米胚中还含有γ-谷维素能降低胆固醇,有利皮肤抗衰老,γ-阿魏酸能抗血栓形成、抗自由基等,N-去氢神经酰胺具有抑制黑色素生成、美化皮肤的作用等。
表3 稻米中已知的活性成分及其功能和对应种质资源

Table 3.Known active ingredients and their functions and corresponding germplasm resources in rice
3.4 胚乳中主要活性物质及其调控机制
胚乳中富含淀粉,抗性淀粉(Resistant Starch,RS)作为一类特殊的淀粉类型,是人体小肠内难以消化吸收的淀粉及淀粉降解物的总称。高抗性淀粉含量的稻米可以增加饱腹感,可有效预防和控制糖尿病,并对肥胖症和肠道疾病起到积极预防作用[37-41]。
Zhou等分离鉴定了稻米抗性淀粉含量调控基因(Soluble starch synthase),其编码可溶性淀粉合酶。该基因突变后显著提高了水稻胚乳中抗性淀粉、直链淀粉、脂类、直链淀粉-脂质复合体含量。进一步研究发现,和(颗粒结合淀粉合酶)共同参与抗性淀粉的合成,对抗性淀粉的调控依赖于基因的高表达,在背景下下调的表达导致抗性淀粉含量下降,直链淀粉-脂质复合体的减少。
进一步研究表明,SSIIIa通过影响对AGPase和PPDK的活性,分别促进直链淀粉和脂类的合成,最终通过形成直链淀粉-脂质复合体提高了胚乳中抗性淀粉含量[41]。高抗性淀粉品种选育也取得了一些进展,如降糖稻1号、宁农黑粳、功米3号、优糖稻2号、优糖稻3号、宜糖1号等[42]。
此外,低谷蛋白含量稻米对于肾病患者具有良好的辅助疗效。已有研究表明,稻米谷蛋白首先在内质网中合成谷蛋白前体,然后通过高尔基体转运到蛋白贮藏囊泡中加工成成熟的谷蛋白。和都参与谷蛋白前体正常转运的各个环节,GPA3能够直接与GPA1/ OsRAB5A/GLUP4和GPA2/OsVPS9A/GLUP6互作,三者形成一个调控复合体,协同调控水稻中致密囊泡介导的后高尔基体谷蛋白前体的运输[43-48]。
上述任何一个组分的突变,均可导致稻米中谷蛋白合成受阻、含量降低。目前,现有的低谷蛋白品种有LGC-1、W3360、W0868、低谷1号和武2812等[49-52]。因此,稻米中生理活性物质合成和代谢途径关键基因的克隆与分子机制解析,为研究和开发功能稻米奠定了理论基础。
此外,中国稻种资源丰富,系统深入地进行生理活性物质的研究与相关种质鉴定,将为我国功能稻米研究提供强有力的材料优势[1]。
4 功能稻米的创制方法
功能稻米的研发途径与常规水稻品种选育方法类似,但是目的和导向性有显著差异。主要包括以下几种形式:一是常规选育与标记辅助结合。利用已有功能稻米种质资源,根据外观、感官表型杂交、回交选育。例如,黑米、紫米、红米和绿米等功能稻米,外观易于辨别,无需其他辅助手段,选择育种效率高。
此外,具有香味的香米种质选育可通过闻、咀嚼等感官判断。此类型功能稻米的选育,是将表型标记与常规选育结合,具有快速、准确、成本低的优点,但也仅限于外观较易辨别的品种类型。有些活性物质或特殊成分并不会引起稻米外观变化,无法用表型标记判断,此时,就需要在明确活性物质合成或代谢关键基因基础上,采用分子功能标记辅助与常规选育相结合,加快选育进程。
二是诱变选育。主要是针对特定的功能成分进行诱变筛选、鉴定,可分为化学诱变、辐射诱变和组培诱变等。例如,在越光遗传背景下通过化学诱变筛选,选育“富含铁”水稻新品种GCN4和系026,对贫血患者效果明显[53];Maeda 等利用金南风经N-甲基-N-亚硝基脲(MNU)化学诱变选育出的“巨大胚”突变体EM40与高产品种Akenohoshi 杂交,培育了首个巨胚稻品种“Haiminori”[33]。
在日本优 (Nihonmasari)遗传背景下,通过辐射诱变选育出了低谷蛋白水稻品种“LGC-1”, LGC-1 稻米中谷蛋白含量低,可吸收的总蛋白明显减少,可供肾病和糖尿病患者食用[52]。以LGC-1为供体亲本,选育出了系列低谷蛋白品种W3360、武2812等[49,51]。
福建农林大学作物遗传育种研究所采用Co60-γ射线辐射诱变常规品种、三系保持系、恢复系,从龙特蒲B、1813B、明恢86等中获得了籼型的巨胚种质,进而育成了TgeB、TgeA、1813geB、1813geA和MgeR86等一批巨胚常规籼稻、巨胚籼型不育系、巨胚籼型恢复系[54]。
中国水稻研究所黄大年等以水稻品种“秀水110”为材料,通过组织培养和系统选育培育出巨胚稻“伽马-1”(基尔-1)等(https:// www.ricedata.cn/variety/varis/608971.htm),并以此品种为基础开发了基尔米、基尔米奶等系列产品。诱变育种主要优点在于筛选出的种质资源及其衍生品种,可直接用于功能稻米改良、生产、推广和食用。但诱变育种也存在诱变产生突变频率较低、工作量大等限制因素。
三是基因工程技术改良。基因工程技术改良是通过对稻米中生理活性物质合成关键基因进行表达修饰,或者对某些胚乳中无法直接合成的活性物质,在胚乳中进行合成途径重构,使其在胚乳中特异积累。例如,通过在水稻中胚乳特异性表达了水仙花来源的八氢番茄红素合成酶和细菌来源的八氢番茄红素去饱和酶,在水稻胚乳中建立了原本没有的β-胡萝卜素合成途径,使得胚乳中可积累β-胡萝卜素,最高可达31.1 μg/g,而β-胡萝卜素作为维生素A 合成的直接前体,用于维生素A的合成[55]。
而虾青素(Astaxanthin)是一类橙红色类胡萝卜色素,为类胡萝卜素的最高级形式,具有超强抗氧化活性,对人体健康具有重要的作用。Zhu等利用水稻胚乳特异性启动子和自主开发的高效多基因TGSII系统,通过分析水稻类胡萝卜素合成途径基因的表达模式,发现大多数类胡萝卜素合成的相关基因在水稻胚乳中处于不表达或低表达状态。
在此基础上,结合对“黄金大米(Golden Rice)”的分析,同时表达八氢番茄红素合成酶基因()、八氢番茄红素脱氢酶基因()、β-胡萝卜素酮化酶基因()和β-胡萝卜素羟化酶基因()四个类胡萝卜素合成途径的关键基因,在水稻胚乳中重新构建了不同基因组合的类胡萝卜素/酮式胡萝卜素/虾青素的生物合成途径,获得富含高抗氧化活性虾青素且转基因筛选标记剔除的虾青素大米“赤晶米”[56]。
同样,叶酸(维生素B9)在胚乳中含量也很低,其由蝶呤、氨基苯甲酸和谷氨酸缩合而成。Storozhenko 等利用水稻胚乳特异性启动子驱动拟南芥中的GTP 环化水解酶Ⅰ(GTP cyclohydrolaseⅠ, GTPCHⅠ,催化蝶呤合成第一步反应)和氨基脱氧分支酸酶(aminodeoxychorismate synthase, ADCS,催化氨基苯甲酸合成第一步反应)编码基因,成功将水稻胚乳中叶酸相对含量提高了15~100倍,叶酸含量最高的转基因稻米中叶酸含量为17.23 μg/g(38.3 nmol/g)[57]。
此外,研究人员在导入和基因的基础上,将经过修饰的、动物来源的叶酸结合蛋白(Folate binding protein,FBP)导入水稻胚乳中来提高稻米中叶酸的稳定性,使得转基因稻米中叶酸的含量进一步提高到对照叶酸含量的150 倍左右,最高达到25.3±1.41 μg/g[58]。
此外,还有γ-氨基丁酸、抗性淀粉、铁、锌、花青素和α-亚麻酸等活性物质或微量元素,通过基因工程手段在水稻胚乳中进行高浓度富集[59-66](表4)。利用基因工程技术可极显著提高胚乳中活性物质含量,然而,按国家现行的转基因作物管理法规,基因工程产品现阶段无法直接种植、食用,稻米中高含量的活性物质需进一步加工提取、提炼制备成次级产品。
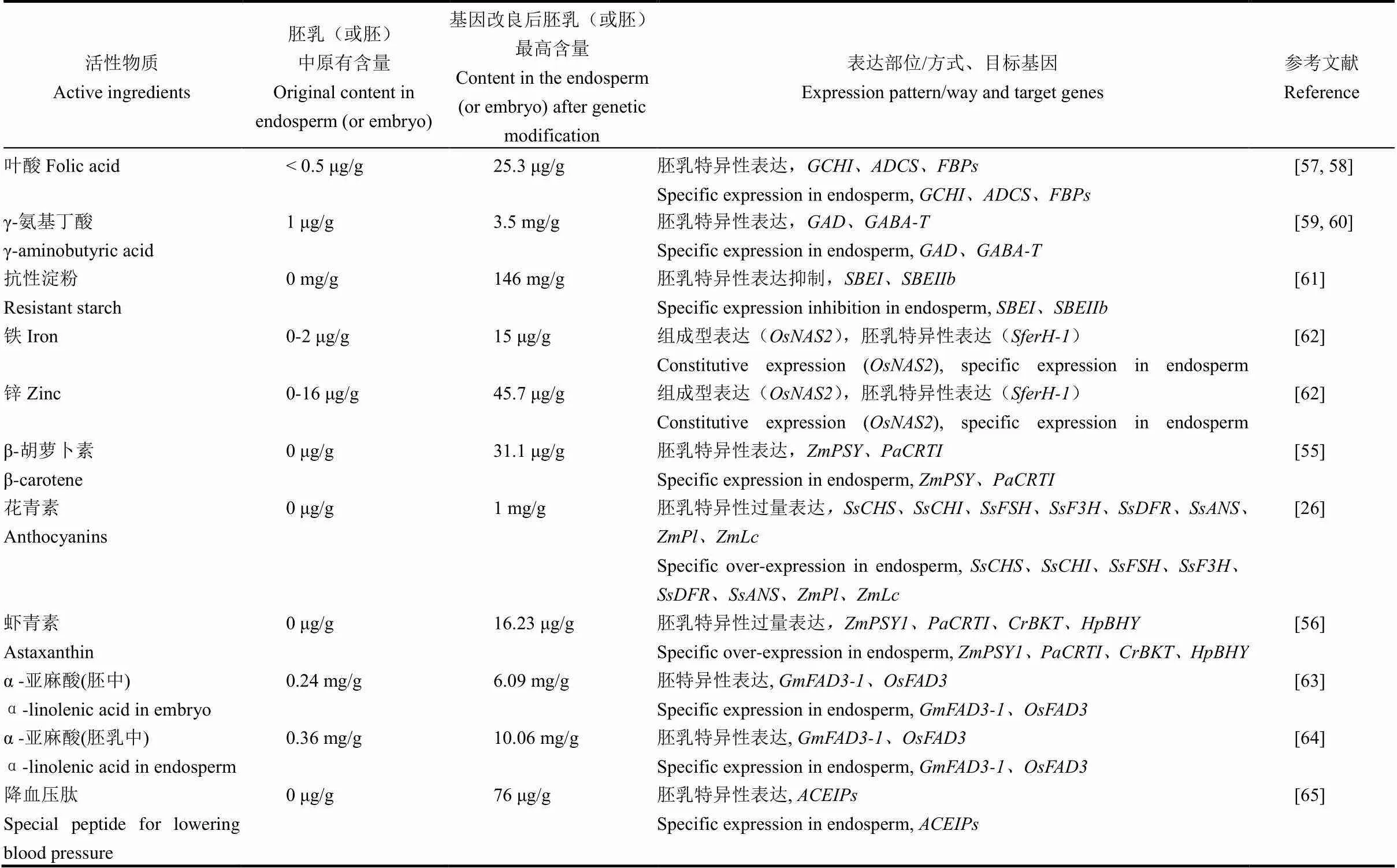
表4 基因工程手段增加稻米中活性物质含量
四是栽培措施强化。通过不同的施肥方式、施肥时期强化活性物质的吸收、转化过程。比如,在灌浆期叶面喷施亚硒酸钠(富硒增产剂),无机态硒经水稻体内生物转化成有机态硒贮藏籽粒中,便于人体有效吸收利用[67, 68]。还可通过调节光照时间和强度、温度等环境因素,促进黑米、紫米及红米中花色素苷的合成和积累[69, 70]。该方法不受种质资源限制,但是受活性物质种类限制,预期效果也较难控制。
五是通过外源物理添加。在稻米加工过程中与外源活性物质进行混合加工工艺处理,例如,将普通大米与按比例混合的维生素B1、维生素B2、烟酸、铁等元素的溶液混合吸胀,再在大米表面喷涂含玉米朊(不溶于水的蛋白)的酒精溶液薄膜,以避免洗淘大米时维生素流失,蒸饭时薄膜融化后营养元素扩散到米饭中[71]。
另外,把提取的降血肽活性物质作为添加剂加入大米中,食用后可降低血压,并增强了免疫系统的功能[71]。虽然将功能稻米研发途径和选育过程进行了分类,但是在实际的选育过程中,要想加快功能稻米研发进程,往往是多种方法和手段相互配合使用,并没有明显的区分,而只是更加侧重于哪种方式进行选育。
5 功能稻米研究中存在的问题及未来发展方向
5.1 功能稻米研究中存在的问题
长期以来,国内水稻育种目标以高产、优质(外观食味为主)、抗病虫等重要农艺性状为主,功能稻米相关研究关注较少且不受重视,导致功能稻米相关研究推进缓慢。通过分析当前功能稻米研究现状、对比国外功能稻米发展形势,当前功能稻米研究主要存在如下急需解决的问题:
一是基础研究薄弱。功能稻米相关的活性物质合成、代谢途径及其调控机理较复杂, 部分活性物质鉴定和分析方法开发成本高。二是功能稻米相关种质资源相对薄弱。种质资源的匮乏,使得种质资源交流变得更加困难,严重阻碍了功能稻米选育的进程。
三是功能稻米的类型有待进一步丰富。现有的功能水稻品种及稻米类型偏少,其选育与利用的系统性有待加强。四是功能稻米推广与产业化乏力。消费者对功能稻米的认知和接受程度远远不足,其产业化延伸力度和深度不够,功能稻米及其衍生产品类型偏少,具有影响力的功能稻米品牌稀缺。
五是功能稻米研究中学科交叉、协作缺乏。功能稻米研究和开发等整个产业涉及遗传育种、营养学、医学、食品科学、分析化学等多个学科以及农业、食品加工、市场营销等多部门,需要多学科、多部门的协作,需要形成大的团队,分工合作,有序推进。六是功能稻米国家或行业标准有待完善。目前为止,国内尚无可参照的功能稻米及其制品国家或行业标准,影响功能稻米产业的健康发展。
5.2 功能稻米研究未来发展方向
基于上述问题及未来发展趋势,功能稻米研究及开发应着力于以下几个方面:一是加强功能稻米相关基础研究与种质创制。功能稻米相关新种质创制,是基础研究与育种应用的重要基础。提高稻米中生理活性物质含量的一个有效途径是发掘富含特定生理活性物质的种质资源,利用传统的育种方法。
结合现代基因突变技术和分子生物学技术,提高稻米中特定生理活性物质含量。加强功能稻米相关基础与应用基础研究,解析稻米中各种生理活性成分代谢有关的基因,促进功能稻米基础研究与育种应用的有效衔接,提高我国功能稻米的科技含量。
同时,要加快与功能稻米相关的各种生理活性成分鉴定和分析方法的建立,加强功能稻米相关生理活性成分加工提取关键技术突破,提升产品的国际竞争力,可有效缓解水稻生产中“天花板”和“地板”困境。
二是加强复合型功能稻米选育和功能稻米药用价值挖掘。中华民族有“药食同源”的传统认知,自唐代以后,在诸多的古代中医药著作中都记述了稻米的药用功能和治疗某些疾病的效果。例如,《黄帝内经·素问》记载“汤液醪醴(稻米五谷制成)护胃气”;李时珍《本草纲目》记载香米能“润心肺、和百药, 久服轻身延年”;孙思邈《千金翼方》记载“常食谷皮(富含VB1),可免脚气病生”。
1911年,荷兰医生艾克曼(Christiaan Eijkman)因从米糠中提取出小分子物质维生素B1,可以治疗脚气病,在1929 年获得诺贝尔生理学与医学奖。以上史书记载表明,人们深刻地认识到稻米或特定稻米类型具有药用性,稻米既能食用、食补又可食治。
因此,根据各种活性物质的功能差异,开展功能稻米或复合型功能稻米的基础研究,通过明确功能稻米中各种活性物质的作用机制,进一步探索和提升功能稻米作为类中药、或在传统中药配方中作为原料或辅料的作用,助力水稻由传统“粮食作物”向“粮食、经济兼顾作物”转型升级,促进功能稻米研究和现代营养医学的共同发展。
三是加强功能稻米的推广与产业化。首先,功能稻米的推广与产业化,离不开有影响力的品牌打造,从而提升功能稻米产品的知名度。目前功能稻米及其衍生制品相关品牌偏少,要通过整合现有稻米品牌资源,根据不同类型功能水稻品种种植具有较强地域性的特点,培育地域性优势或强势品牌,坚持走“少、精、尖”的品牌战略之路,通过特有功能和品质打响品牌,取信于市场,不断增强消费者的信心。
其次,功能稻米产品的多元化有利于提升功能稻米产业化的广度,从初级米制品(适糖米、宜糖米)到米胚油、米糠油,到以功能稻米为原材料的食品添加剂、糕点、饮料等深加工产品[72],将提升功能稻米的附加值,拓宽功能稻米产业化路子。
第三,将胚乳作为生物反应器,通过活性成分在胚乳中特异表达、纯化,提升功能稻米产业化的深度。武汉大学研究团队等利用水稻胚乳细胞特异高效表达技术及活性蛋白纯化工艺,先后表达纯化了重组人血清白蛋白(OsrHSA)、重组人碱性成纤维生长因子(OsrbFGF)、重组人抗胰蛋白酶(OsrAAT)、重组人表皮生长因子(OsrEGF)、重组人乳铁蛋白(OsrLF)和重组人类胰岛素生长因子(OsrIGF-1)等[73-76],部分产品直接进入医疗临床试验或进一步用于医药制品、保健品和美容护肤品的生产制备。
例如,人血清白蛋白(HAS)作为人血浆中含量最丰富的蛋白,是激素、脂类等物质的转运载体,其主要生理功能是调节血浆pH值和维持血浆渗透压。而OsrHSA是一种来源于转基因水稻胚乳的重组人血清白蛋白,不含有动物源成分,能为无血清培养基提供更安全的选择。
与胎牛血清(FBS)、血浆来源白蛋白(pHSA)和牛血清白蛋白(BSA)相比,OsrHSA具有更高的纯度,更好的批次稳定性,可以更深入地进行临床研究[71]。因此,通过以水稻胚乳为生物反应器,极大提升了功能稻米的附加值,促进了功能稻米产业链的延伸。
四是加强功能稻米大众化消费的宣传和舆论引导。功能稻米不仅是针对于“病人”,也不仅仅有疾病辅助治疗的功能,其还具有良好的疾病预防功效,适合所有人群。而现有稻米作为主食的消费习惯和大众对功能稻米认知的局限,使得绝大多数人群对功能稻米的作用产生误解,认为只有生病了才有必要去选择功能稻米替代常规稻米。
因此,正确有效的宣传和舆论引导必不可少。首先,要从市场和消费者的角度对功能稻米及制品相关的专业术语,进行规范、统一。在已有的文献报道中,有功能性稻米、功能型稻米、功能稻米、营养大米、功能大米和功能稻等不同的称谓,还有一些根据稻米中成分含量来命名的富硒米、富铁米等。
从专业的角度来认识和理解这些不同称谓的含义难度不大,但是从面向消费者的角度考量,上述不同的称谓势必会对消费者造成困扰。因此,规范和统一具有特殊功能的稻米及其制品的学术称谓,对提高“功能稻米”的行业辨识度、正确引导消费者具有重要意义。
同时,结合当前“健康中国行动(2019-2030年)”等国家战略实施的契机,在功能稻米相关研究基础上,推广“营养健康型稻米”概念和相关品牌,消除普通大众对功能稻米只适合“病人”的误解,将“营养健康型稻米”理念、产品引入大众日常饮食中,优化居民的健康饮食结构,逐渐提升消费者对功能稻米及其产品的黏性,将有利于功能稻米及其衍生产品的可持续发展。
此外,制定和完善功能稻米及其制品国家标准,将营养功能品质作为优质稻米的标准之一,加强功能稻米产学研一体化、上中下游的有机衔接,加强多学科、多部门的分工协作等,必将大力促进功能稻米产业化的健康发展,也必将促进稻米生产加工由“满足消费者需求”向“引领消费者需求”的转换升级。
[1] 胡培松.功能性稻米研究与开发[J].中国稻米, 2003, 5: 3-5.
Hu P S.Research and development of functional rice [J]., 2003, 5: 3-5.(in Chinese)
[2] 樊胜根.全球背景下的中国粮食安全与营养[J].中国发展观察,2015(1): 87-92.
Fan S G.Food security and nutrition in China in the global context [J].2015(1): 87-92.(in Chinese)
[3] 苏宁, 万向元, 翟虎渠, 万建民.功能型水稻研究现状和发展趋向[J].中国农业科学, 2007, 40(3): 433-439.
Su N, Wan X Y, Zhai H Q, Wan J M.Progress and prospect of functional rice researches[J]., 2007, 40(3): 433-439.(in Chinese with English abstract)
[4] 卢士军, 黄家章, 吴鸣, 沈东婧, 孙君茂.营养导向型农业的概念、发展与启示[J].中国农业科学, 2019, 52(18): 3083-3088.
Lu S, Huang J Z, Wu M, Shen D J, Sun J M.The concept and development of nutrition-sensitive agriculture and its enlightenments to China[J]., 2019, 52(18): 3083-3088.(in Chinese with English abstract)
[5] Serageldin I.Biotechnology and food security in the 21st century[J]., 1999, 285: 387-391.
[6] Potrykus I.Golden rice and beyond [J]., 2001, 125: 1157-1161.
[7] 周惠明, 张民平.糙米中功能性成分的研究[J].食品科技, 2002(5): 17-19.
Zhou H M, Zhang M P.Research on functional components in Brown rice [J]., 2002(5): 17-19.(in Chinese)
[8] 江友兴, 刘希贞.黑米成分分析与功能利用[J].农牧产品开发, 1999(8): 8-9.
Jiang Y X, Liu X Z.Composition analysis and Functional Utilization of black rice [J]., 1999(8): 8-9.(in Chinese)
[9] OECD.Safety assessment of foods and feeds derived from transgenic crops, common bean, rice, cowpea and apple compositional considerations [M],, 2019, 3.WHO, “Energy and protein requirements”, in Joint FAO/ WHO/ UNU Expert Consultation on Protein and Amino Acid Requirements in Human Nutrition., 2007, No.935.
[10] 孙定红.我国功能性稻米及其食品的研究进展[J].粮食加工, 2006, 31(5): 20-23.
Sun D H.Research progress of functional rice and its food in China [J]., 2006, 31(5): 20-23.(in Chinese)
[11] 顾华孝.米糠的食用性和在保健功能食品上的应用[J].粮食与饲料工业, 2001(5): 46-48.
Gu H X.The eating of rice bran and its application in functional nutritious food [J]., 2001(5): 46-48.(in Chinese)
[12] 徐浩.米糠资源研究与应用[D].合肥: 安徽农业大学, 2015.
Xu H.Research and application of rice bran resources [D].Hefei: Anhui Agricultural University, 2015.(in Chinese with English abstract)
[13] Satake T.Modern rice milling technology [M].Tokyo, Japan: University of Tokyo Press, 1990: 295.
[14] 陈浩.功能性水稻研究思路和前景分析[J].生命科学, 2016, 28 (10): 1279-1286.
Chen H.Research methods and prospects of functional rice [J]., 2016, 28 (10): 1279-1286.(in Chinese with English abstract)
[15] 张艳华.巨胚稻生物学特征及营养成分研究[D].福州: 福建农林大学, 2010.
Zhang Y H.Study on biological characteristics and nutritional components of giant embryo rice [D].Fuzhou: Fujian Agriculture and Forestry University, 2010.(in Chinese with English abstract)
[16] 黄大年, 钱前, 郭龙彪, 颜美仙.稻米的营养和人类健康.中国稻米, 2008(6): 16-19.
Huang D N, Qian Q, Guo L B, Yan M X.Nutrition of rice and human health., 2008(6): 16-19.(in Chinese)
[17] 赵其国, 尹雪斌, 孙敏, 刘永贤, 侯非凡, 张宁.2008-2018 年功能农业的理论发展与实践[J].土壤, 2018, 50(6): 1061-1071.
Zhao Q G, Yin X B, Sun M, Liu Y X, Hou F F, Zhang N.A ten-year overview of functional agriculture from 2008 to 2018 [J]., 2018, 50(6): 1061-1071. (in Chinese with English abstract)
[18] 赵其国, 尹雪斌.功能农业[M].北京: 科学出版社, 2016.
Zhao Q G, Yin X B.Functional agriculture [M].Beijing: Science Press, 2016.(in Chinese)
[19] Hu J, Anderson B, Wessler S R.Isolation and characterization of rice R genes: Evidence for distinct evolutionary paths in rice and maize[J]., 1996, 142: 1021-1021.
[20] Hu J, Reddy V S, Wessler S R.The ricegene family: Two distinct subfamilies containing several miniature invertedrepeat transposable elements[J]., 2000, 42: 667-678.
[21] Sakamoto W, Ohmori T, Kageyama K, Miyazaki C, Saito A, Murata M, Noda K, Maekawa M.The purple leaf () locus of rice: Theallele has a complex organization and includes two genes encoding basic helix-loop- helix proteins involved in anthocyanin biosynthesis[J]., 2001, 42: 982-991.
[22] Rahman M M, Lee K E, Lee E S, Matin M N, Lee D S, Yun J S, Kim J B, Kang S G.The genetic constitutions of complementary genesanddetermine the purple color variation in pericarps with cyanidin-3-O-glucoside depositions in black rice[J]., 2013, 56: 24-31.
[23] 王彩霞, 舒庆尧.水稻紫色种皮基因的精细定位与候选基因分析[J].科学通报, 2007, 52(21): 2517-2523.
Wang C X, Shu Q Y.Fine localization and candidate gene analysis ofin purple seed coat of rice [J]., 2007, 52(21): 2517-2523.(in Chinese)
[24] 庄杰云, 杨长登, 钱惠荣, 赵成章, 郑康乐.紫米基因与RFLP标记的连锁分析[J].遗传学报, 1996, 23(5): 372-375.
Zhuang J Y, Yang C D, Qian H R, Zhao C Z, Zheng K L.Linkage analysis of purple rice gene and RFLP marker [J]., 1996, 23(5): 372-375.(in Chinese with English abstract)
[25] Oikawa T, Maedab H, Oguchia T, Yamaguchi T, Tanabe N, Ebana K, Yano M, Ebitani T, Izawa T.The birth of a black rice gene and its local spread by introgression[J]., 2015, 27: 2401-2414.
[26] Zhu Q L, Yu S Z, Zeng D C, Liu H M, Wang H C, Yang Z F, Xie X R, Shen RX, Tan J T, Li H Y, Zhao X C, Zhang Q Y, Chen Y L, Guo J X, Chen L T, Liu Y G.Development of ‘‘Purple Endosperm Rice’’ by engineering anthocyanin biosynthesis in the endosperm with a high-efficiency transgene stacking system[J]., 2017, 10: 918-929.
[27] Furukawa T, Maekawa M, Oki T, Suda I, Iida S, Shimada H, Takamure I, Kadowaki K.Theandgenes are involved in proanthocyanidin synthesis in rice pericarp[J]., 2006, 49(1): 91-102.
[28] Gu X Y, Foley M E, Horvath D P, Anderson J, Feng J H, Hang L H, Mowry C R, Ye H, Suttle J C, Kadowaki K, Chen Z X.Association between seed dormancy and pericarp color is controlled by a pleiotropic gene that regulates abscisic acid and flavonoid synthesis in weedy red rice[J]., 2011, 189(4): 1515-1524.
[29] Sweeney M T, Thomson M J, Pfeil B E, McCouch S R.Caught red-handed:encodes a basic helix-loop-helix protein conditioning red pericarp in rice [J]., 2006, 18: 283-294.
[30] Sweeney M T, Thomson M J, Cho Y G, Park Y J, Williamson S H, Bustamante C D, McCouch S R.Global dissemination of a single mutation conferring white pericarp in rice[J]., 2007, 3(8): 1418-1424.
[31] 李勇.发芽糙米的储藏特性及其GABA含量测定方法的研究[D].成都: 西华大学, 2014.
Li Y.Study on the storage characterication of germinated brown rice and the determination method of its GABA content [D].Chengdu: Xihua University, 2014.(in Chinese with English abstract)
[32] 刘玲珑, 江玲, 刘世家, 周时荣, 张文伟, 王春明, 陈亮明, 翟虎渠, 万建民.巨胚水稻W025糙米浸水后γ-氨基丁酸含量变化的研究[J].作物学报, 2005, 10: 1265-1270.
Liu L L, Jiang L, Liu S J, Zhou S R, Zhang W W, Wang C M, Chen L M, Zhai H Q, Wan J M.Accumulation of the γ-aminobutyric acid content in brown rice: A new rice strain W025 with giant-embryo during water soaking [J]., 2005, 10: 1265-1270.(in Chinese with English abstract)
[33] Maeda H, Nemoto H, Iida S.A new rice variety with giant embryos, “Haiminori” [J]., 2001, 51: 211-213.
[34] Xu F, Fang J, Ou S J, Gao S P, Zhang F X, Du L, Xiao Y H, Wang H R, Sun X H, Chu J F, Wang G D, Chu C C.Variations incoding region influence grain size and yield in rice[J]., 2015, 38(4): 800-811.
[35] Nagasawa N, Hibara K, Heppard E P, Vander Velden K A, Luck S, Beatty M, Nagato Y, Sakai H.encodes CYP78A13, required for proper size balance between embryo and endosperm in rice[J]., 2013, 75(4): 592-605.
[36] Yang W B, Gao M J, Yin X, Liu J Y, Xu Y H, Zeng L J, Li Q, Zhang S B, Wang J M, Zhang X M, He Z H.Control of rice embryo development, shoot apical meristem maintenance, and grain yield by a novel cytochrome P450[J]., 2013, 6(6): 1945-1960.
[37] 张虹, 杨晓飞, 李倩.高抗性淀粉大米对2型糖尿病病人餐后血糖波动的影响[J].全科护理, 2019, 17(24): 3008-3010.
Zhang H, Yang X F, Li Q.Effect of high resistant starch rice on postprandial blood glucose fluctuation in type 2 diabetic patients[J]., 2019, 17(24): 3008-3010.(in Chinese)
[38] 曾亚文, 杨树明, 杜娟, 吴殿星, 普晓英, 房亚南.高抗性淀粉稻米防治慢性病研究进展[J].农业科技通讯, 2009, 1: 37-39.
Zeng Y W, Yang S M, Du J, Wu D X, Pu X Y, Fang Y N.Research progress in the prevention and treatment of chronic diseases in rice with high resistant starch [J]., 2009, 1: 37-39.(in Chinese)
[39] 吴伟, 刘鑫, 杨朝柱, 舒小丽, 吴殿星.抗性淀粉及预防糖尿病和肥胖症功能稻米研究进展[J].核农学报, 2006, 20(1): 60-63.
Wu W, Liu X, Yang C Z, Shu X L, Wu D X.Progress in the study of resistant starch and function rice of for prevention of diabetes and hyperlipidemia., 2006, 20(1): 60-63.(in Chinese with English abstract)
[40] 方长云, 胡贤巧, 卢林, 段彬伍.稻米抗性淀粉的研究进展[J].核农学报, 2015, 29(3): 513-520.
Fang C Y, Hu X Q, Lu L, Duan B W.Advances in Study on rice resistant starch[J]., 2015, 29(3): 513-520.(in Chinese with English abstract)
[41] Zhou H J, Wang L J, Liu G F, Meng X B, Jing Y H, Shu X L, Kong X L, Sun J, Yu H, Smith M S, Wu D X, Li J Y.Critical roles of soluble starch synthase SSIIIa and granule-bound starch synthase Waxy in synthesizing resistant starch in rice[J]., 2016, 113(45): 12844-12849.
[42] 王瑞智, 张得雯, 王娜, 田蕾, 杨淑琴, 李培富.宁夏高抗性淀粉水稻种质资源筛选及粒形相关性分析[J].种子, 2015, 34(9): 6-10.
Wang R Z, Zhang D W, Wang N, Tian L, Yang S Q, Li P F.The selection of high resistant starch content of rice germplasm resources in Ningxia and correlation analysis of grain shape [J]., 2015, 34(9): 6-10.(in Chinese with English abstract)
[43] Fukuda M, Satoh-Cruz M, Wen L, Crofts A, Sugino A, Washida H, Okita T, Ogawa M, Kawagoe Y, Maeshima M, Kumamaru T.The small GTPase Rab5a is essential for intracellular transport of proglutelin from the Golgi apparatus to the protein storage vacuole and endosomal membrane organization in developing rice endosperm [J]., 2011, 157(2): 632-644.
[44] Fukuda M, Wen L, Satoh-Cruz M, Kawagoe Y, Nagamura Y, Okita T, Washida H, Sugino A, Ishino S, Ishino Y, Ogawa M, Sunada M, Ueda T, Kumamaru T.A guanine nucleotide exchange factor for Rab5 proteins is essential for intracellular transport of the proglutelin from the Golgi apparatus to the protein storage vacuole in rice endosperm [J]., 2013, 162(2): 663-674.
[45] Liu F, Ren Y L, Wang Y H, Peng C, Zhou K N, Lv J, Guo X P, Zhang X, Zhong M S, Zhao S L, Jiang L, Wang H Y, Bao Y Q, Wan J M.OsVPS9A functions cooperatively with OsRAB5A to regulate post-Golgi dense vesicle-mediated storage protein trafficking to the protein storage vacuole in rice endosperm cells [J]., 2013, 6(6): 1918-1932.
[46] Ren Y L, Wang Y H, Liu F, Zhou K N, Ding Y, Zhou F, Wang Y, Liu K, Gan L, Ma W W, Han X H, Zhang X, Guo X P, Wu F Q, Cheng Z J, Wang J L, Lei C L, Lin Q B, Jiang L, Wu C Y, Bao Y Q, Wang H Y, Wan J M.encodes a regulator of post-Golgi vesicular traffic essential for vacuolar protein sorting in rice endosperm [J]., 2014, 26(1): 410-425.
[47] Wang Y H, Ren Y L, Liu X, Jiang L, Chen L M, Han X H, Jin M N, Liu S J, Liu F, Lv J, Zhou K N, Su N, Bao Y Q, Wan J M.regulates endomembrane organization and storage protein trafficking in rice endosperm cells [J]., 2010, 64(5): 812-824.
[48] Wen L Y, Fukuda M, Sunada M, Ishino S, Ishino Y, Okita T, Ogawa M, Ueda T, Kumamaru T.Guanine nucleotide exchange factor 2 for Rab5 proteins coordinated with GLUP6/GEF regulates the intracellular transport of the proglutelin from the Golgi apparatus to the protein storage vacuole in rice endosperm [J]., 2015, 66(20): 6137-6147.
[49] 吴越, 胡静, 江祺祥, 刘古春, 杨明方, 徐晓杰.功能性专用水稻品种武2812的选育[J].浙江农业科学, 2011(6): 1323-1324.
Wu Y, Hu J, Jiang Q X, Liu G C, Yang M F, Xu X J.Breeding of functional special rice variety Wu 2812 [J]., 2011(6): 1323-1324.(in Chinese)
[50] Iida S, Amano E, Nishio T.A rice (L.) mutant having a low content of glutelin and a high content of prolamine [J]., 1993, 87(3): 374-378.
[51] 蔡金洋, 杨尧城, 徐伟东, 李白, 李军.利用分子标记辅助选育低谷蛋白水稻株系[J].浙江农业学报, 2015, 27(9): 1505-1509.
Cai J Y, Yang Y C, Xu W D, Li B, Li J.Breeding of rice lines with low glutelin content by molecular marker-assisted selection[J]., 2015, 27(9): 1505-1509.(in Chinese with English abstract)
[52] Kusaba M, Miyahara K, Iida S, Fukuoka H, Takano T, Sassa H, Nishimura M, Nishio T.: A dominant mutation that suppresses the glutelin multigene family via RNA silencing in rice[J]., 2003, 15(6): 1455-1467.
[53] 胡繁荣, 段智英, 张琳琳, 胡培松, 吴殿星.稻米功能性成分育种研究进展[J].核农学报, 2004, 18(5): 364-367.
Hu F R, Duan Z Y, Zhang L L, Hu P S, Wu D X.Advances in rice breeding for the functional components [J]., 2004, 18(5): 364-367.(in Chinese with English abstract)
[54] 杨艳荔.优质巨胚稻营养成分分析及发芽试验[D].福建: 福建农林大学, 2008.
Yang Y L.Nutritional composition analysis and germination test of high-quality giant embryo rice [D].Fujian: Fujian Agriculture and Forestry University, 2008.(in Chinese with English abstract)
[55] Paine J A, Shipton C A, Chaggar S, Howells R M, Kennedy M J, Vernon G, Wright S Y, Hinchliffe E, Adams J L, Silverstone A L, Drake R.Improving the nutritional value of golden Rice through increased pro-vitamin A content [J]., 2005, 23(4): 482-487.
[56] Zhu Q L, Zeng D C, Yu S Z, Cui C J, Li J M, Li H Y, Chen J Y, Zhang R Z, Zhao X C, Chen L T, Liu Y G.From golden rice to aSTARice: Bioengineering astaxanthin biosynthesis in rice endosperm [J]., 2018, 11: 1440-1448.
[57] Storozhenko S, Brouwer V D, Volckaert M, Navarrete O, Blancquaert D, Zhang G F, Lambert W, Straeten D V D.Folate fortification of rice by metabolic engineering [J]., 2007, 25: 1277-1279.
[58] Blancquaert D, Van Daele J, Strobbe S, Kiekens F, Storozhenko S, Steur H D, Gellynck X, Lambert W, Stove C, Straeten D V D.Improving folate (vitamin B9) stability in biofortified rice through metabolic engineering [J]., 2015, 33: 1076-1078.
[59] Shimajiri Y, Oonishi T, Ozaki K, Kainou K, Akama K.Genetic manipulation of the gamma-aminobutyric acid (GABA) shunt in rice: Overexpression of truncated glutamate decarboxylase (GAD2) and knockdown of gamma-aminobutyric acid transaminase (GABA-T) lead to sustained and high levels of GABA accumulation in rice kernels [J]., 2013, 11: 594-604.
[60] Akama K, Kanetou J, Shimosaki S, Kawakami K, Tsuchikura S, Takaiwa F.Seed-specific expression of truncated OsGAD2 produces GABA-enriched rice grains that influence a decrease in blood pressure in spontaneously hypertensive rats [J]., 2009, 18: 865-876.
[61] Zhu L J, Gu M H, Meng X L, Cheung S C K, Yu H X, Huang J, Sun Y, Shi Y C, Liu Q Q.High-amylose rice improves indices of animal health in normal and diabetic rats [J]., 2012, 10(3): 353-362.
[62] Trijatmiko K R, Dueñas C, Tsakirpaloglou N, Torrizo L, Arines F M, Adeva C, Balindong J, Oliva N, Sapasap M V, Borrero J, Rey J, Francisco P, Nelson A, Nakanishi H, Lombi E, Tako E, Glahn R P, Stangoulis J, Chadha-Mohanty P, Johnson A A T, Tohme J, Barry G, Slamet-Loedin I H.Biofortified indica rice attains iron and zinc nutrition dietary targets in the field., 2016, 6: 19792.
[63] Yin Z J, Liu H L, Dong X B, Tian L H, Xiao L, Xu Y N, Qu L Q.Increasing α-linolenic acid content in rice bran by embryo-specific expression of ω3/△15-desaturase gene [J]., 2014, 33: 987-996.
[64] Liu H L, Yin Z J, Xiao L, Xu Y N, Qu L Q.Identification and evaluation of ω-3 fatty acid desaturase genes for hyperfortifying α-linolenic acid in transgenic rice seed [J]., 2012, 63(8): 3279-3287.
[65] Qian D D, Qiu B, Zhou N, Takaiwa F, Yong W D, Qu L Q.Hypotensive activity of transgenic rice seed accumulating multiple antihypertensive peptides [J]., 2020, 68(27): 7162-7168.
[66] Zhao M C, Lin Y J, Chen H.Improving nutritional quality of rice for human health [J]., 2020, 133: 1397-1413.
[67] 郭天宇.叶面喷施不同硒肥对水稻含硒量、产量及品质的影响[D].哈尔滨: 东北农业大学, 2016.
Guo T Y.Effects of different foliar selenium fertilizers on the selenium content, yield and quality of rice [D].Harbin: Northeast Agricultural University, 2016.(in Chinese with English abstract)
[68] 段门俊, 田玉聪, 吴芸紫, 金涛, 陈阜, 刘章勇.叶面喷施亚硒酸钠对再生稻产量及品质的影响[J].中国水稻科学, 2018, 32(1): 96-102.
Duan M J, Tian Y C, Wu Y Z, Jin T, Chen F, Liu Z Y.Effect of foliar application of Na2SeO3on the yield and quality of ratooning rice [J]., 2018, 32(1): 96-102.(in Chinese with English abstract)
[69] 柳敏, 王忠, 顾蕴洁.有色稻颖果的发育和色素沉积[J].中国水稻科学, 2011, 25(4): 392-398.
Liu M, Wang Z, Guo Y J.Caryopsis development and anthocyanidin accumulation of colored rice [J]., 2011, 25(4): 392-398.(in Chinese with English abstract)
[70] 蔡光泽.环境因素对有色米糙米着色程度的影响蔡光泽[J].中国农学通报, 2003, 19(4): 71-74.
Cai G Z.Influence of applying color the degree of the environment factor to the color rice brown rice[J]., 2003, 19(4): 71-74.(in Chinese with English abstract)
[71] 黄大年, 钱前, 郭龙彪, 颜美仙.功能性稻米的加工途径及其前景 [J].农产品加工, 2009, 3: 8-9.
Huang D N, Qian Q, Guo L B, Yan M X.Processing methods and prospects of functional rice [J]., 2009, 3: 8-9.(in Chinese)
[72] 杨春华, 杨明毅, 刘刚.发芽糙米食品产业化的开发[J].粮油食品科技, 2003, 11(2): 12-13.
Yang C H, Yang M Y, Liu G.Development of germinated brown rice food industrialization[J]., 2003, 11(2): 12-13.(in Chinese)
[73] He Y, Ning T T, Xie T T, Qiu Q C, Zhang L P, Sun Y F, Jiang D M, Fu K, Yin F, Zhang W J, Shen L, Wang H, Li J J, Lin Q S, Sun Y X, Li H Z, Zhu Y G, Yang D C.Large-scale production of functional human serum albumin from transgenic rice seeds [J].2011, 108 (47): 19078-19083.
[74] An N, Ou J Q, Jiang D M, Zhang L P, Liu J R, Fu K, Dai Y, Yang D C.Expression of a functional recombinant human basic fibroblast growth factor from transgenic rice seeds., 2013, 14: 3556-3567.
[75] Zhang L P, Shi J N, Jiang D M, Stupak J, Ou J Q, Qiu Q C, An N, Li J J, Yang D C.Expression and characterization of recombinant human alpha-antitrypsin in transgenic rice seed., 2012, 164: 300-308.
[76] Luo J L, Ning T T, Sun Y F, Zhu J H, Zhu Y G, Lin Q S, Yang D C.Proteomic analysis of rice endosperm cells in response to expression of hGM-CSF., 2009, 8: 829-837.
国稻种芯百团计划行动 胡时开-胡培松:功能性稻米现状与展望
 热门关注
热门关注 毛主席长叹一声!林彪叛逃后家中搜出一物
毛主席长叹一声!林彪叛逃后家中搜出一物

